
An overview of microglia subtypes and markers
Related
Microglia are the immune effector cells of the central nervous system (CNS) existing in three distinct forms which serve different functional roles. Find out more about these cell types and how to use markers to identify them.
Amoeboid microglia
Amoeboid microglia are associated with the developing CNS. In rats, amoeboid microglia have been shown to appear late in gestation and disappear soon after birth1,2.
These cells exhibit a round cell body, possess pseudopodia and thin filopodia-like processes and contain numerous lysosomes; all traits indicative of a motile phagocytic phenotype.
During the post-natal period, amoeboid microglia are believed to play a role in tissue histogenesis through the removal of inappropriate and superfluous axons3,4 and through the promotion of axonal migration and growth5.
Ultimately, amoeboid microglia grow long crenulated processes and transform into ramified microglia found in the adult CNS6,7.
Ramified microglia
Ramified microglia are present in abundance in the brain parenchyma and constitute approximately 10–20% of the total population of glial cells in the adult8,9.
These small round cells comprise numerous branching processes and possess little cytoplasm. In the adult brain, the resident population of ramified microglia is maintained through local cell division and through the recruitment of circulating peripheral blood monocytes10.
Under physiological conditions, microglia exhibit pinocytotic activity and localized motility11–13. It has been suggested that ramified microglia contribute to metabolite removal and to the clearance of toxic factors released from injured neurons13.
A study has demonstrated that microglia have the propensity to transform into neurons, astrocytes or oligodendrocytes14. Hence, ramified microglia may represent a unique population of multipotent stem cells in the adult CNS, which strongly implicates microglia in CNS repair.
Learn more about the functions of glia in the CNS.
Reactive microglia
In response to injury or pathogen invasion, quiescent ramified microglia proliferate and transform into active 'brain macrophages' otherwise known as reactive microglia15,16.
Microglial proliferation can be studied using antibodies raised against nuclear antigens such as Ki6717,18 or proliferating cell nuclear antigen (PCNA)19 or by bromodeoxyuridine (BrdU) staining20.
Reactive microglia are rod-like, devoid of branching processes and contain numerous lysosomes and phagosomes. The reactive cell form represents a population of macrophages, which are associated with brain injury and neuroinflammation.
Following a damaging event, reactive microglia accumulate at the site of injury18,21,22 where they play a neuroprotective role phagocytosing damaged cells and debris.
In acute lesions the peak of microglial activation occurs 2-3 days post insult, but if the pathological stimulus persists microglial activation continues9.
Reactive microglia express MHC class II antigens and other surface molecules necessary for antigen presentation including CD40, B7 and ICAM-123,24. Consequently, microglia are considered to be the most potent antigen-presenting cells in the CNS. Like macrophages, reactive microglia secrete inflammatory mediators, which orchestrate the cerebral immune response.
Chronic microglial activation is associated with neurological disorders including Alzheimer's disease25,26, multiple sclerosis27,28 and delayed neuronal death occurring after ischaemia29,30.
In these instances, the persistent activation of microglia accompanied by the sustained secretion of inflammatory mediators is thought to have a deleterious effect on neuronal function and survival, thereby exacerbating disease processes.
Identification of microglia using antigenic markers
A paucity of specific microglial-only antigens has hindered microglial identification. Markers used for the detection of microglia are also present in macrophage since both cell types exhibit the same lineage.
Microglia are often identified using flow cytometry as it enables to quantify the differences in antigen expression levels. Ramified parenchymal microglia have been demonstrated to possess the phenotype CD11b+, CD45low, whilst other CNS macrophages and peripheral macrophages exhibit the phenotype CD11b+, CD45high (Figures 1 and 2)31,32.

Figure 1: Expression of CD45 by microglia.
CD45 expression by microglia (mic) extracted from 5-day old rat neonates as previously described33,34. Microglia were isolated and left in culture for 24 hours.
The cells were subsequently harvested, fixed, then analyzed by flow cytometry using anti-CD45 (ab8216). The labeled cells are represented by the black shaded populations, whereas the unlabeled cells are depicted by the grey line (%: % of cells in M1 or M2 region, MFI: mean fluorescence intensity).

Figure 2: Expression of CD45 by peritoneal macrophages
Extraction and staining were performed as in Figure 1. Labeled cells are represented by the black shaded populations, whereas the unlabeled cells are depicted by the grey line.
Microglia can also be detected immunologically using antibodies raised against macrophage-specific antigens; however, such antibodies fail to distinguish microglia from macrophages.
The OX-42 antibody33,35 (illustrated in Figure 3) recognizes the CR3 complement receptor (CD11b/CD18) expressed by rat or mouse microglia. Clone F4/80 binds a 60 kDa glycoprotein on murine ramified microglia36.
Alternatively, CD68 is a lysosomal protein and can be used to stain microglia37,38 as shown in Figure 4. High levels of CD68 expression are associated with macrophages (Figure 5) and activated microglia, while low levels of expression are associated with quiescent ramified microglia33,37,38.
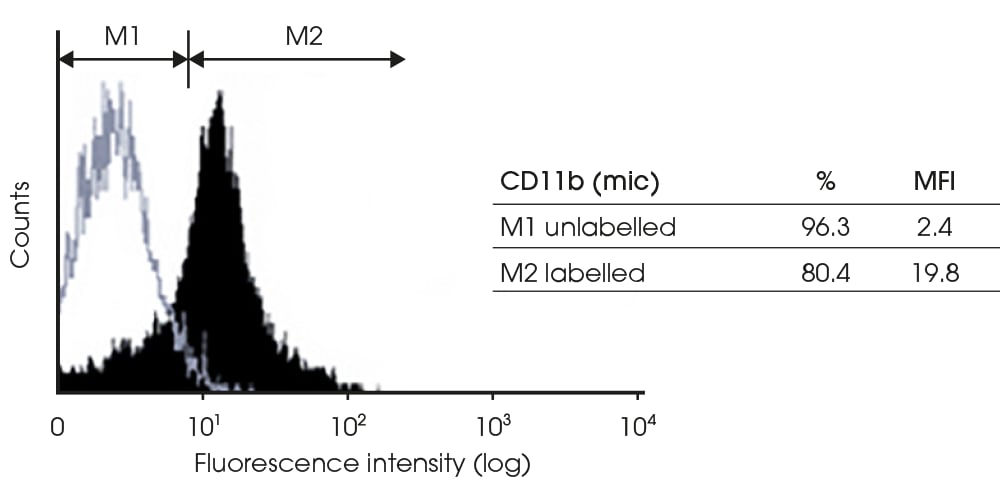
Figure 3: Expression of CD11b by microglia
CD11b expression by microglia (mic) extracted from 5-day old rat neonates as previously described33,34.
Microglia were isolated and left in culture for 24 hours. The cells were subsequently harvested, fixed, then analyzed by flow cytometry using the OX-42 antibody. Labeled cells are represented by the black shaded populations, whereas unlabeled cells are depicted by the grey line (%: % of cells in M1 or M2 region, MFI: mean fluorescence intensity).

Figure 4: Expression of CD68 by microglia
CD68 expression by microglia (mic). Cells were extracted from 5-day old rat neonates as previously described33,34.
Microglia were isolated and left in culture for 24 hours. The cells were subsequently harvested, fixed then analyzed by flow cytometry using anti-CD68 (ED-1) antibodies. Labeled cells are represented by the black shaded populations, whereas the unlabeled cells are depicted by the grey line (%: % of cells in M1 or M2 region, MFI: mean fluorescence intensity).

Figure 5: Expression of CD68 by peritoneal macrophages
Identification of microglia can also be achieved using Rio Hortega's original silver carbonate staining technique39 or using lectin staining40. Lectins are carbohydrate-binding proteins that label microglia through the recognition of glycoproteins containing terminal alpha-D-galactose residues41.
Furthermore, microglia cells can be identified using antibodies raised against the intermediate protein vimentin42,43 or by using acetylated low-density lipoprotein (LDL) conjugated to a fluorescent tag, which enables the labeling of LDL receptors44,45.
Learn more about glia
The functions of glia in the CNS
Radial glia cell markers and their major functions
The role of glia in demyelinating diseases pathway
References
Written by Claudie Hooper and Jennifer Pocock.
J.M. Pocock is at Cell Signalling Laboratory, Department of Neuroinflammation, Institute of Neurology, University College London, 1 Wakefield Street, London, WC1N 1PJ.